近日,Plant Biotechnology Journal杂志在线发表了由扬州大学焦新安和刘巧泉课题组合作撰写的“Rice-derived SARS-CoV-2 glycoprotein S1 subunit vaccine elicits humoral and cellular immune responses”论文。该研究工作通过构建含不同启动子的载体并转化水稻,获得表达新冠病毒S1蛋白的转基因水稻。rS1蛋白可在转基因水稻种子中表达,且S1基因插入不影响水稻籽粒大小和重量。将纯化的rS1蛋白与铝佐剂联合使用,可诱导小鼠产生增强的体液免疫应答,与鼠伤寒沙门菌鞭毛蛋白(FliC)佐剂联合使用,具有诱导体液和细胞免疫潜力。研究团队报道的这一基于水稻生物反应器表达亚单位疫苗策略,为开发安全、有效且经济的新冠疫苗提供了新路径,并且这一策略可以普遍应用于其它病毒疫苗的研发。

新冠病毒(SARS-CoV-2)自 2019 年以来在全球肆虐,尽管已有多种商业疫苗获批,但病毒持续传播与变异,疫情防控形势依旧严峻。据世界卫生组织报道,截至2024年8月,全球累计确诊超7.75亿例,死亡超700万例。开发能诱导有效免疫应答、安全且经济的疫苗,成为全球尤其是发展中国家的迫切需求。
SARS-CoV-2的S蛋白在病毒入侵宿主细胞过程中起关键作用,其S1亚基上的RBD结构域与人体ACE2受体结合,介导病毒感染,因此S1亚基成为疫苗研发的核心靶点。前期研究探索了多种疫苗策略,亚单位疫苗因安全性高而备受关注,植物作为生产疫苗抗原的平台,以其成本低、易规模化、无病原体污染及能进行真核蛋白修饰等优势,展现出巨大潜力。全文主要研究结果如下:
1.构建水稻表达系统并鉴定rS1蛋白表达
科研人员构建了两个含SARS-CoV-2 S1基因的二元载体,pCAMBIA1300Gt1-S1(水稻谷蛋白启动子Gt1)和 pCAMBIA1300Actin-S1(组成型启动子Actin)(图1a),并通过农杆菌介导转化水稻品种 ZH11,最终获得 56 株独立的T0代阳性转基因水稻植株。随后,科研人员对后代进行筛选。利用PCR分析技术,检测水稻植株中是否含有目标基因;借助潮霉素抗性分离分析,确定T3-1至T3-6植株为纯合子。利用Western blot和质谱分析技术,科研人员在pGt1::S1转基因水稻种子中检测到rS1蛋白。分子量约110 kDa的rS1蛋白条带。质谱分析确认该蛋白为rS1蛋白,检测到S1氨基酸序列的51%,且经PNGase F处理后,rS1蛋白分子量变为约76 kDa,表明发生了N-糖基化修饰(图1b-e)。与之形成对比的是,pActin::S1转基因株系未检测到rS1蛋白表达,凸显了pGt1启动子在驱动rS1蛋白表达方面的优势。
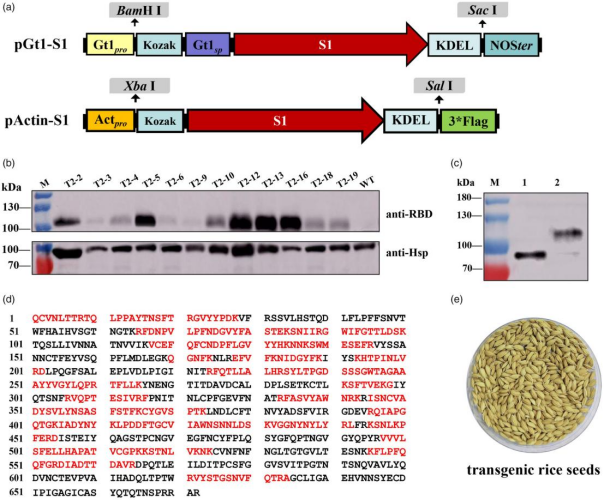
图1 水稻表达载体构建及rS1蛋白表达鉴定
2.rS1蛋白的定量与种子农艺性状
研究团队采用基于探针的qRT-PCR技术,对纯合水稻株系(T3代)中rS1蛋白的转录水平进行了测定。通过已知浓度的含S1质粒生成标准曲线,计算出 S1基因的拷贝数。在6个纯合株系中,T3-6转基因植株种子中的S1基因拷贝数最高(图2a)。利用定量Western blot分析测定rS1蛋白在纯合水稻株系中的表达水平,结果显示T3-6转基因水稻株系的表达水平最高,达到282 μg/g干重(图2b),这与qRT- PCR分析结果一致。基于这些结果,研究团队选择T3-6转基因水稻株系进行后续深入研究。为评估S1基因表达对水稻种子的影响,研究团队对T3代转基因植株(1-6号株系)的种子进行了性状考察。结果显示,转基因植株种子的粒长、粒宽和千粒重(TGW)与野生型对照相比,均无显著差异(图2c)。这一发现表明,S1基因的插入并未对水稻种子的大小和重量产生影响,为后续利用水稻生产疫苗提供了重要的可行性依据。
3.rS1蛋白的纯化与ACE2结合活性
研究人员选取表达水平最高的T3-6转基因水稻种子进行蛋白纯化。结果显示,纯化后的rS1蛋白分子量约为110 kDa,且该蛋白与兔抗RBD蛋白多克隆抗体表现出强烈且特异的反应性,表明其具有良好的免疫反应性(图2d, e)。通过ELISA分析S1蛋白与ACE2蛋白的结合活性,发现S1蛋白与ACE2的结合亲和力随ACE2 浓度的增加而增强。水稻来源的rS1(rS1 glyc)与HEK293细胞产生的S1相比,结合力较弱,但在ACE2浓度升高至100 μg/mL时,其结合亲和力接近HEK293产生的S1。此外,在较低ACE2浓度(<50 μg/mL)下,去糖基化的rS1表现出更强的结合力,而在较高浓度(>50 μg/mL)下,糖基化的rS1结合效果更佳(图2f)。糖基化与去糖基化的rS1蛋白展现出不同结合优势,为理解rS1蛋白生物学特性和疫苗优化设计提供了关键线索。
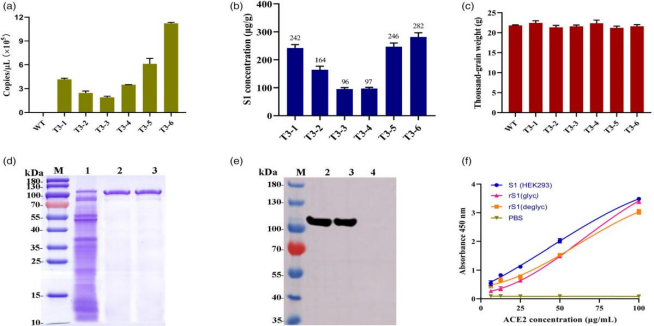
图2 rS1蛋白的定量、纯化及ACE2结合活性
4.rS1疫苗诱导体液免疫应答
为评估rS1的免疫原性,研究团队以BALB/c小鼠为实验对象,进行了免疫实验。分别肌肉注射三次rS1蛋白,同时设置了添加FliC佐剂、Alum佐剂以及不添加佐剂的组别,每次免疫间隔2周(图3a)。在第二次或第三次接种2周后,发现rS1免疫组小鼠血清中S1特异性IgG抗体滴度显著高于PBS对照组。而且,添加FliC和Alum佐剂的rS1免疫组,其IgG抗体水平显著高于未添加佐剂的rS1免疫组。第三次接种后,rS1诱导产生的S1特异性IgG滴度相较于第二次接种后提高了10倍(图3b)。此外,Alum佐剂的rS1免疫组诱导产生了更高的IgG1抗体反应,而FliC佐剂的rS1疫苗则显著提高了IgG1和IgG2a抗体反应(图3c)。为进一步探究rS1疫苗产生的抗体效果,研究人员进行SARS-CoV-2假病毒进行中和试验。结果显示,rS1 + FliC和rS1 + Alum组小鼠血清中的中和抗体(NAb)滴度显著高于rS1单独免疫组(图3d)。这表明添加佐剂的rS1疫苗能够有效中和SARS-CoV-2假病毒,为其作为疫苗的有效性提供了有力证据。
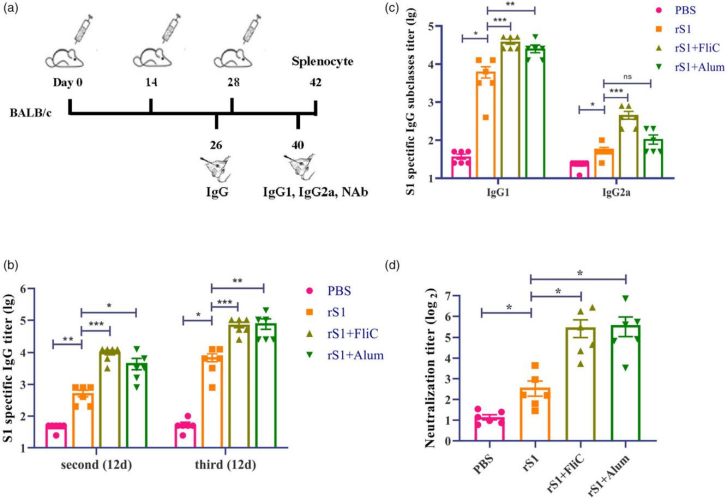
图3 rS1疫苗诱导体液免疫应答
5.rS1疫苗诱导细胞免疫应答
在第三次接种2周后,研究团队采集小鼠脾脏,通过细胞增殖和细胞因子表达分析来评估疫苗诱导的细胞免疫应答。qRT-PCR检测结果显示,与PBS组相比,rS1组小鼠体内Th1细胞因子(IFN-γ)、Th2细胞因子(IL-4和IL-10)以及Th17细胞因子(IL-17A)的mRNA表达水平显著增加。当rS1与Alum联合使用时,TNF-α、IL-4和IL-10的表达水平显著升高,而IFN-γ和IL-17A的表达无显著差异。当rS1与FliC联合使用时,IFN-γ、TNF-α、IL-4、IL-10和IL-17A的表达均显著高于rS1单独组(图4a)。此外,通过ELISA检测脾细胞在rS1蛋白刺激两天后的细胞因子分泌水平,得到了与qRT-PCR分析一致的结果(图4b)。同时,rS1 + FliC组的脾淋巴细胞增殖水平显著高于rS1 + Alum组(图4c)。ELISpot结果显示rS1组中分泌IFN-γ和IL-4的脾淋巴细胞数量显著高于PBS组,S1 + FliC免疫组中分泌IFN-γ和IL-4的脾淋巴细胞数量显著高于rS1免疫组,表明FliC佐剂增强了rS1抗原特异的Th1/Th2混合型免疫应答。S1 + Alum免疫组中分泌IL-4的脾淋巴细胞数量显著高于rS1免疫组,而分泌IFN-γ的淋巴细胞数量与rS1免疫组相比无显著差异,表明Alum佐剂增强了抗原特异的Th2免疫应答(图 4d)。
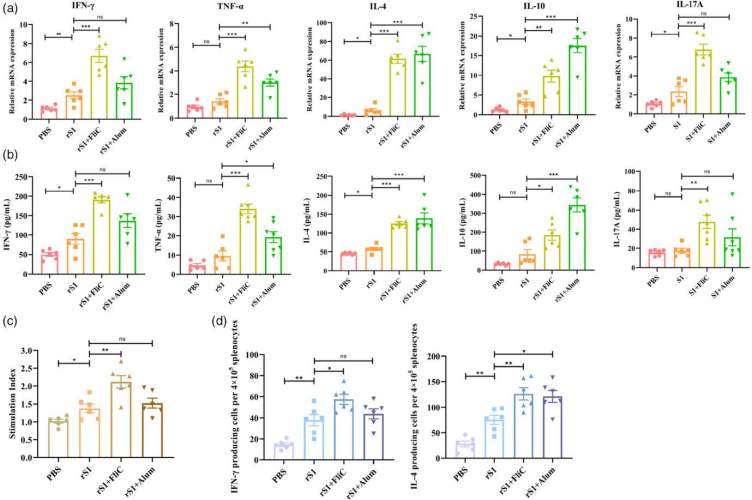
图4 rS1疫苗诱导细胞免疫应答
6.rS1疫苗诱导细胞因子分泌的CD4+和CD8+ T细胞免疫应答
通过流式细胞术对细胞因子分泌的CD4+和CD8+ T细胞进行定量分析。与仅使用rS1免疫相比,rS1联合鞭毛蛋白FliC进行免疫接种,使得分泌IFN-γ和IL-4的CD4+ T细胞的比例更高,同时分泌IL-17A的CD4+ T细胞的表达量也有所增加。然而,在rS1 + Alum组中,只有分泌IL-4的CD4+ T细胞显著增加(图5a)。在rS1+FliC组中,观察到抗原特异性的分泌IFN-γ的CD8+ T细胞的比例显著增加。各组之间分泌IL-4的CD8+ T细胞和分泌IL-17A的CD8+ T细胞的比例没有显著差异(图5b)。
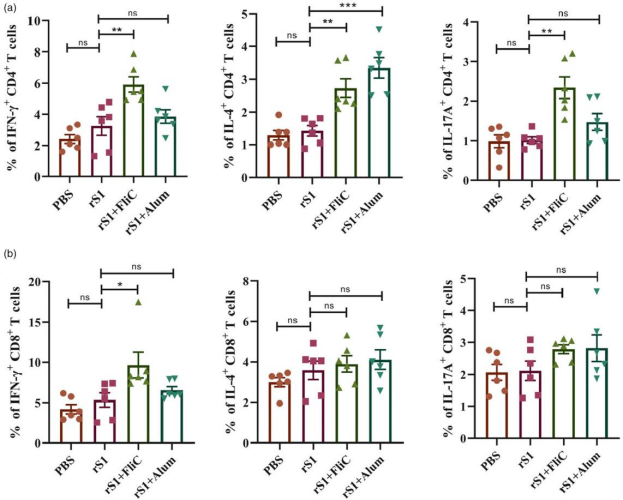
图5 rS1疫苗诱导CD4+和CD8+ T细胞免疫应答
这项研究研发的基于水稻表达系统的新冠S1亚单位疫苗策略,不仅展示了其在诱导体液免疫应答和细胞免疫应答中的有效性,也揭示了水稻作为疫苗生产平台的巨大潜力。鞭毛蛋白FliC佐剂促进了混合型Th1/Th2/Th17免疫,为优化疫苗配方、提高疫苗免疫效果提供了重要的实验依据。本研究为深入开发新冠疫苗及应对其它病毒病提供了新路径。
扬州大学宋丽讲师、硕士生温亚亚为论文共同第一作者,扬州大学焦新安教授、刘巧泉教授、潘志明教授为该研究工作共同通讯作者。研究工作得到了国家重点研发计划(2022YFC2604200)、国家自然科学基金(32102679)、江苏省重点研发计划(现代农业)(BE2021331)等项目的资助。
论文链接:
https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.70077
(原文来源于植物生物技术Pbj公众号推文)

 苏公网安备 32100302010246号
苏公网安备 32100302010246号